|
����2�����
����2.1������״̬���������7�����õ�˫������״̬�����Բ��졣��60�������ߣ���t���飬�������������֮������и߶�������ͳ��ѧ���壨P��0.001����������֮�������������ͳ��ѧ���壨P��0.05�������1��
��1���ξ��������������ȱ仯����D��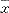 ��s�� ��s��
| ��� |
ʵ���� |
������ |
| A�� |
+0.65��1.64 |
+3.35��1.05 |
| B�� |
+0.72��1.23 |
+3.15��0.98 |
| C�� |
+0.69��0.85 |
+3.75��0.73 |
| ����2.2���۳��������������60��������������2��
��2���������������ѧ��������mm����s�� |
| ��� |
�۱� |
ǰ����� |
������ |
������ǻ���� |
���᳤�� |
������ǻ����
���������᳤�� |
| A�� |
�� |
2.50��0.37 |
4.95��0.19 |
7.94��0.65 |
15.39��0.65 |
0.52��0.03 |
| �� |
2.56��0.35 |
4.74��0.59 |
7.21��0.79** |
14.53��0.72** |
0.48��0.02** |
| B�� |
�� |
2.80��0.14 |
4.92��0.17 |
7.45��0.47 |
15.01��0.53 |
0.50��0.02 |
| �� |
2.76��0.14 |
4.97��0.23 |
6.94��0.32** |
14.78��0.70** |
0.47��0.01** |
| C�� |
�� |
2.85��0.22 |
4.99��0.25 |
7.29��0.55 |
15.14��0.68 |
0.49��0.02 |
| �� |
2.77��0.12 |
5.03��0.20 |
6.83��0.41** |
14.63��0.62** |
0.46��0.02** |
���������۱Ƚϣ�*��*P��0.01
����2.3������ѧ�۲�
����2.3.1���⾵������ʵ���۵Ĺ�Ĥ��ԭ��ά���䱡�����Ρ���ϸ����״����֯�ۼ�����Ƭ״������Ĥ������ĤҲ�䱡���ԡ�����������Բ��졣
����2.3.2����羵������3��
��3��������羵�۲�
| �� |
��Ĥ |
��״�� |
����Ĥ |
| ������ |
��ԭ��ά�ʿ�״��Բ��״��������ֱ��45mm��ͼ1�� |
��ɫ����Ƥϸ������зḻ�����壬ɫ��ϸ������������ɫ�ؿ��� |
ɫ����Ƥ����Χ����ɫ�أ��ɼ���ڡ��ڽڡ��ں˲㼰����ά��ͼ4�� |
| A�� |
��ԭ��ά����״��ֱ��3mm��ͼ2�� |
��ɫ����Ƥϸ������ߣ��˺��ѷ��֣�ɫ��ϸ������Χɫ�ؿ������࣬��Ĥ��������������Ⱦɫ�������״��ϸ�������Ըı� |
ɫ����Ƥ����Χɫ���������٣������ṹ��֯δ�����Ըı䣨ͼ5�� |
| B�� |
��ԭ��ά�ı�ͬA�飬ֱ��Ϊ4mm��ͼ3�� |
��Ĥ���ݱ��ԣ�Һ�������ܽ�ϸ���ʣ���������ʧ |
ͬA�飨ͼ6�� |
| C�� |
ͬA�� |
ͬA�� |
ͬA�� |
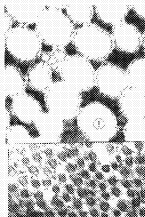


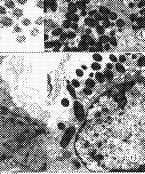

ͼ1�������۹�Ĥ��ԭ��ά�����棬ֱ��Լ45mm����5000
2��A��ʵ���۹�Ĥ��ԭ��ά�����棬ֱ��Լ3mm����5000
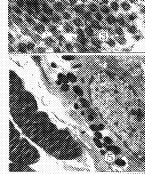
3��B��ʵ���۹�Ĥ��ԭ��ά�����棬ֱ��Լ4mm����5000
4������������Ĥɫ����Ƥ�㣬�����д���ɫ�ء���5000
5��A��ʵ��������Ĥɫ����Ƥ�㣬����ɫ�ش������١���5000
6��B��ʵ��������Ĥɫ����Ƥ�㣬��A�����ơ���5000
����3������
����3.1�����ξ��������ģ��
����ͨ���ξ����᷽�����о����������ʵ�鶯��缦[1]��������[2]��è[3]����[4]�ȵİ����Խ���ģ�ͣ���ӿ�������Խ��ӵ���ʶ���̣�ͬʱҲΪҩ����ƽ����ṩʵ�������������ݡ���ʵ��ѡ���ã���Ȼ��������Ĥ�й�ϸ���ֲ��������нϴ���죬����ʵ��Ƕȿ�������һ����ֵ��ͬʱҲ�Ⱥ��ʵ�鶯�ᆳ�õöࡣĿǰ�������ð����Խ���ʵ�鱨�����������ߵ���������������ԭ������У����ñ�����ĸ��ι�����ɻ��ʵͣ��˹�ι���������ܳɻ���۷����ڳ�������ȱ仯���������Խ�����Ҫ�ϳ����ڸ�ʱ�䣬��Ҫ�ڳ�ʱ����ʹ������ȫ�պ��Ҳ���ŧ��Ⱦ���dz����ѣ����Ա�ʵ����ڸ���53�죬ʵ���۵������Ϊ+0.65��1.64D��δ�����Խ��ӣ��������+3.35��1.05D��ȣ�ֻ��˵�ǡ���������Խ��ӡ�������ǰ��ʵ������һ���IJ�ͬ����������������Ƶ�A��֤ʵ�ξ������ܵ����۵������ӳ������в�����ǻ�ӳ��Ͳ�����ǻ���ȣ����᳤��ռ������λ����ij�̶ֳ���˵��������ǻ���ȣ����᳤�ȱȲ�����ǻ���ȸ��ܷ�ӳ������ǻ�ڽ��ӷ����е����á������������ɰ����Խ��ӵ���Ҫ��̬ѧԭ���䲡��ѧԭ��֮һ�ǹ�Ĥ��ԭ��ά�ı�ϸ�ͱ䳤�����ڹ⾵�͵羵�о��õ�֤ʵ���羵�²�������۹�Ĥ��ԭ��άֱ����45nm����A����3nm������15������һ��ԭ�����������Ĥɫ����Ƥ��ı䡣�ڵ羵������Ĥ�������ṹ��δ���ı䡣
����3.2���ȵ��غͽ��ӵĹ�ϵ
����JoshWallman[5]���������Ĥ���ƽ������ۣ���Ϊ��Ĥ������һ������ԭ������Ϣ����ֱ��Ӱ�칮Ĥ��������Щ��Ϣ�����ܴ�����Ĥɫ����Ƥ��Ľ������ϣ�������Ĥ��Ѫ�ܵ�����ڵĹ�Ĥ����һ�������ܵ�ԭ�����źż�������Ĥ�źŲ���ɫ����Ƥ�źţ������յ����ڹ�Ĥ����������Ĥ�źš��ڱ�ʵ��������Ĥ������Ĥ�����������ڵ羵�¿ɼ�����Ĥɫ����Ƥ��ı䣬��Щ��֧������Ĥ�������ۡ�Ŀǰ����ܵ�����Ĥ�����ź��Ƕ�Ͱ�[5��6��7]��Ͱ���������������ܽ��ͼ�����Ȱ����Խ����۵������ӳ�����һ�����ܵ��ź����ȵ���[5]��Ȼ���ڱ�ʵ�����ȵ��ز�������ֹ�����Խ��ӵķ�������չ������ȵ��ؿ��ܲ�������Ĥ�����źš���֮��������Ϊ���ξ������ܵ�������������ӳ���������ǻ�ӳ�������̬ѧ��ԭ��Ĥ��ԭ��ά��ϸ���Σ�����Ĥ������Ĥ�������䲡��ѧԭ���ξ������ƻ�������Ĥ������Ĥ���ϣ���������Ĥ�Թ�Ĥ������ʧ�أ��Ӷ��������ӣ�Ȼ���ȵ��ز�������ֹ�����Խ��ӵķ�������չ��
���������
����[1]������������£�������. �ξ�����Խ������γɵ�Ӱ��ʵ���о�. �л��ۿ���־��1996��32��61
����[2]��Neville A McBrien��Hadio Moghaddam��Robert New��et al. Experimental myopia in a diurnal mammal��Sciurus Carolinensis��with no accommodative ability. J Physio��Lond����1993��469��427
����[3]��������. ���۷������èʵ���Խ����о�. �ۿ��½�չ��1997��17��68
����[4]��Richard A. Stone��Alan M. Laties��Elio Raviola��et al. Increase in retinal vasoactive intestinal polypeptide after eyelid fusion in primates. Proc Natl Acad Sci USA��1988��85��257
����[5]��Josh. Wallman. Retinal control of eye growth and refraction in progress in retinal research. Great Britain��Pergamon Press Ltd��1993. 133��153
����[6]��Ф�֣�����Ӣ��������. ����������ƽ��ӻ������о�. �л��ۿ���־��1998��34��130
����[7]��Richard A. Stone��Ton Lin��Alan M. Laties��et al. Retinal dopamine and form��deprivation myopia. Proc Natl Acad Sci USA��1989��86��704��706
���ո壺1998��10��15���أ�1999��05��04�� ��һҳ [1] [2] |



