|
3 ����
N e t r i n s ��һ������ϵͳ���������о�����ϸ����ͻ�����ϸ��Ǩ���յ����õķ����͵��ס��� j��������о�����N e t r i n s �����е�N e t r i n - 1 ������һ����Ҫ�Ĵ�Ѫ����������⋯ �� 0��������������Ѫ����Ƥϸ����Ѫ��ƽ����ϸ���о����֣�N e t r i n - 1 ������Ѫ����Ƥϸ����Ѫ��ƽ����ϸ�������������ã� ͬʱ����������ϸ������˿����ԭ�� N e t r i n һ1 �ܴٽ���������������������Ƥϸ������Ѫ����Ƥϸ����Ѫ��ƽ����ϸ��Ǩ�ƺ�����һj����һ���Ļ���ʵ��֤������Դ��N e t r i n �� l ��ֱ���յ�����Ѫ�ܵķ����� ��������ë����Ĥʵ���У� N e t r i n һ1 ��������������ë����Ĥ����Ѫ�ܵķ��������⣬ ���������Ķ�����������֫ȱѪ�� ͨ���ֲ�ע��N e t r i n һ1������������֫ȱѪ���Ĺ�ע�� ��֯ѧ�о������� ȱѪ��Ѫ����ע������������N e t H n һ1�յ������˴���������Ѫ�ܡ�ֵ��һ����ǣ� N e t r i n һ1 �յ�����ë����Ĥ����ȱѪ��֫Ѫ��������������Ѫ����Ƥ��������( v a s c u l a re n d o t h e l i a l g r o w t h f a c t o r �� V E G F ) �൱����Դ��N e t r i n һ1ͬ�����յ���������Ѫ�ܵķ����� ������N e t r i n һ1 ��
VEGF�Ļ��Ϳ���ֲ����ĤԵ1 mm �Ľ�Ĥ��䣬�����ɻ��Ϳ����ͷŵ�Netrin��1���յ���Ĥ����Ѫ�ܵ�
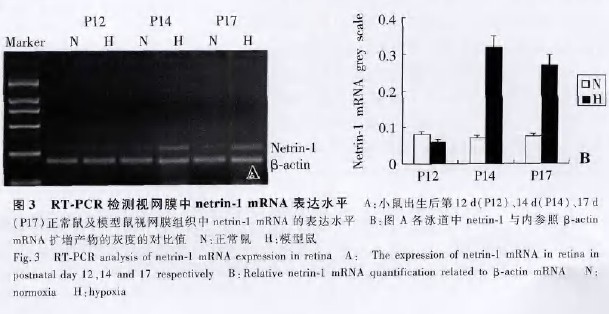
�����������յ���Ĥ����Ѫ�ܵ�������VEGF�൱ ���ɴ˿ɼ�����Դ��Netrinһ1�ܹ�ͨ���ٽ���Ƥϸ����������Ǩ�ƣ�ֱ���յ�����Ѫ�ܵ��γɣ�������ȱѪ��ȱ���Ȳ����̼�������Դ�Դ�Ѫ������������VEGF���Эͬ����������Ѫ�ܵķ��������ڴ����Dz��ø����յ�������Ĥ����Ѫ��ģ�������о���Դ�Ե�Netrinһ1������Ĥ����Ѫ���γɵĵ������á������յ�������Ĥ����Ѫ��С��ģ����1994����Smith�� �ṩ��һ�����ƵĽ�������Ĥ����Ѫ�ܶ���ģ�͵ķ�������ģ�ͷ���������У��ظ��Ժã�����Ѫ�ܵķ�����Ϊ100�� ��Ŀǰ�Ѿ���Ϊ�о�����Ĥ����Ѫ����õĶ���ģ��֮һ�������յ�������Ĥ����Ѫ��ģ�������ó������7 d��С������ĤѪ����δ�������죬����5 d�ĸ��������ڻ���Ը����ı����Է�Ӧ������ĤѪ���ձ��������̶�����ĤѪ�ܹ㷺�������������������к�����Ѫ�ܲ�����Ӧ����Ĥ��֯��������������Ĥ�ڲ���֯���ֳ�����ȱѪ��ȱ����λ������Ĥ�ڲ������Ĥ��ϸ�����ں˲�����Ĥϸ����ȱѪ��ȱ���Ȳ����̼��ºϳɲ��ͷŶ��ִ�Ѫ���������� ����VEGF ���ٺ�ϸ��������(erythropoietin��EPO) �ȣ��Ӷ��ٽ�����Ĥ����Ѫ�ܵ��γɡ����о�����������������12 dʱ��ģ��������Ĥ��֯��Netrinһ1 mRNA����ˮƽ������������Ƚϲ�����ͳ��ѧ���塣���������14 dʱ�������պ�2 d��ģ����������Ĥ��֯��Netrinһ1 mRNA
����ˮƽ�϶����������ϵ������������17 dʱ�������ֺ�5 d��ģ�����ʱ����Ĥ����Ѫ���γ���Ŀ��
�࣬ģ��������Ĥ��֯��Netrinһ1 mRNA����ˮƽ�Ը��ڶ������ϳ���14 dʱNetrin��l mRNA����ˮ
ƽ�������½��������ƶ�ģ��������Ĥ����Ѫ�ܷ��������У�����ȱ��������Ĥ��֯���ܴ�ת¼ˮƽ����
Netrinһ1�ı���Ӷ��յ�����Ĥ����Ѫ�ܵķ���������������ģ��������Ĥ����Ѫ�ܷ��������У�����Ĥ��֯ͨ������Netrinһ1��ת¼�Ӷ��ϵ�Netrinһ1 mRNA����ˮƽ����������Ĥ����Ѫ�ܵ��γɹ���
��Netrin I���ı���仯�����Ƿ���mRNA�仯������һ�£�ȱѪ��ȱ��������Ĥ��֯��ͨ������;����
����Netrinһ1��ת¼�Լ�Netrin-1��ͨ���������弰�ź�;�����ٽ�����Ĥ����Ѫ���γɣ����д��ڽ�һ�����о���
��һҳ [1] [2] [3] [4] [5] [6] ��һҳ |



